Лекция миология филогенез онтогенез и функциональная анатомия мышечной. Филогенез систем органов хордовых Филогенез скелетных мышц
Мышечная система развивается параллельно с развитием нервной системы как ее исполнительный орган. В филогенезе она появляется на основе свойств сократимости элементов клеток простейших многоклеточных организмов, отвечающих на раздражение. Вначале в организме животных появляется гладкая мышечная ткань, еще хорошо сохраняющая клеточное строение и связанная с нервной системой через два нейрона. (Эфферентная связь с центрами нервной системой осуществляется через периферический нейрон.) С совершенствованием форм поступательного движения и связанных с ним опорных элементов тела - скелета появляется скелетная (соматическая) мышечная ткань, у которой уже осуществляется прямая эфферентная связь с центральной нервной системой.
Если у кишечнополостных животных мышечные элементы еще не обособлены, а представлены специальными мышечными отростками эпителиальных клеток, которые дифференцируются на продольно расположенные (в эктодерме) и кольцеобразно (в эндодерме), то у червей они обособляются, уже не связаны с эпителием и образуют кожно-мышечный мешок.
У хордовых, более подвижных животных, все мышцы разделяются на соматические и висцеральные. У взрослых особей соматические мышцы исчерченные. Они представлены продольным пластом парной боковой мышцы, поделенной перепонками на отдельные метамеры (сегменты). В сосудах и кишечнике у ланцетника по-прежнему остается гладкая мышечная ткань. У позвоночных животных, как и у ланцетника, мышцы делятся на соматические и гладкие, различающиеся не только строением, но и характером нервных связей.
Чем сложнее в филогенезе становится организм позвоночных животных, тем больше скелетные мышцы теряют метамерность, тем более дифференцированными они становятся, разделяясь вначале на дорсальные и вентральные пласты, а затем на отдельные слои, отделы, группы мышц и мышцы.
У наземных позвоночных все скелетные мышцы подразделяются уже на группы: мышцы туловища, головы и конечностей. Гладкая мышечная ткань по-прежнему остается главным образом в стенках сосудов и внутренних органов. Как бы ни была сложна скелетная мышца как орган, она связана с определенными сегментами мозга.
Эта связь устанавливается с первых моментов появления мышечных элемен-тов и не нарушается при филогенетических преобразованиях, В связи с этим было установлено - группа мышечных волокон, получающих нервные окон-чания (нервные бляшки) от одной нервной клетки, является «мышечной еди-ницей». Из какого сегмента развивается мышца у эмбриона, из того же сег-мента она иннервируется в дальнейшем. Если в процессе развития мышца перемещается, за ней следует и нерв (яркий пример этого - диафрагма, расположенная позади грудного мозга). Если мышца дифференцируется на отдельные части, нерв подвергается той же дифференцировке; если мышца сливается, сливаются и нервы. Редукция мышцы приводит к редукции и нервной связи. Такая неизменная связь позволяет выявить пути развития мышцы в эмбриогенезе и филогенезе, дает возможность установить, за счет каких сегментов зародыша она сформировалась.
Рис 91. Схема наружных мышц позвоночных
В процессе индивидуального развития (в онтогенезе) мышцы развиваются из нескольких источников мезодермы; 1) за счет материала миотомов мезодермы; 2) из несегментированной мезодермы в области жабер. Поэтому мышцы, развивающиеся за счет мезенхимы жаберного аппарата, называют висцеральными. Вначале у зародыша закладка мышечных элементов, как и в филогенезе, сегментирована и дифференцируется на отдельные метамеры, разделенные соединительнотканными перегородками.
В онтогенезе повторяются основные этапы, особенно раннего периода филогенеза (рис. 91). Вначале боковая мышца закладывается вдоль тела как продольный парный тяж, сегментированный соединительнотканными перегородками на метамерно расположенные миотомы, и каждый миотом получает уже нерв определенного нервного сегмента. Затем появляется продольная соединительнотканная перегородка, разделяющая миотом на дорсальную и вентральную части. В дальнейшем на месте этой продольной перегородки развиваются поперечные отростки позвонков и ребер. В такой же последовательности идет разделение и нервных ветвей, сохраняющих связь с теми же нервными сегментами. За счет материала дорсальных частей миотомов в дальнейшем развиваются мышцы позвоночного столба, а за счет вентральных - мышцы боковых грудных и брюшных стенок.
В зачатки конечностей несколько позже врастают мышечные пучки от дорсальных и вентральных участков миотомов (от пяти миотомов в грудных и от восьми - в тазовых конечностях). Дифференциацией мышечных пластов на отдельные мышцы скелетная мускулатура еще более усложняется, особенно с выделением мышц конечностей, иннервируемых тоже главным образом вентральными ветвями спинномозговых нервов.
Мышцы головы и некоторые мышцы шеи развиваются из несегментированной мезодермы области пяти первых жаберных дуг. Из зачатков первой жаберной дуги развиваются жевательные мышцы, связанные с V парой черепных нервов, из второй - мимические, связанные с VII парой черепных нервов.
Мышечная ткань благодаря свойству сокращаться в процессе филогенеза сформировалась в отдельные органы - мышцы - лентообразные двигатели, обладающие силой, способной поднимать большие тяжести, удерживать на ногах массу тела, наносить удары, развивать большую скорость при передвижении и т. д.
Мышцы, сокращаясь, не только вызывают движение (выполняют динамическую работу), но и обеспечивают тонус мышц, укрепляя суставы под определенным углом сочетания при неподвижном теле, сохраняя определенную позу (выполняют статическую работу). Статическая работа, особенно у крупных (тяжелых) копытных животных, чрезвычайно трудна.
Определенный тонус мышц выполняет и антигравитационную функцию. Вместе с сухожилиями, фасциями и связками скелета мышцы обладают свойствами упругости, благодаря которым являются надежными амортизаторами, придающими конечностям большую рессорность, а также работают как микронасосы, способствующие выведению крови и лимфы из органов (Н. И. Ариинчин, 1987).
У каждого вида животного в процессе эволюции мышцы, развиваясь, определяют свои пределы силы, скорости движения, выносливости, а также и массы, которая находится в прямой зависимости от вида, возраста, пола, породы животного и их тренировки. Масса мышц составляет у крупного рогатого скота и лошадей примерно 42-47% от общей массы тела, у овец 34, а у свиней 31%. Только работа (тренировка) мышц способствует наращиванию их массы как за счёт увеличения диаметра мышечных волокон (гипертрофия), так и за счет увеличения их количества (гиперплазия).
Работа мышц теснейшим образом связана с органом равновесия и в значительной мере с другими органами чувств. Благодаря этой связи мышцы обеспечивают равновесие тела, точность движений, силу. Мышцы - мощный генератор перехода химической энергии пищи в тепловую энергию, энергии упругих деформаций в механическую, использующуюся для продвижения крови по сосудам и возбуждения афферентных импульсов, посылаемых в мозг, а также перехода в тепловую энергию.
Не изолированная мышечная система
Единый кожно-мускульный мешок
Появление поперечно-полосатой мышечной ткани
Разделение мышечных тяжей на миотомы
Развитие групп мышц
Развитие мышц конечностей (изменение среды обитания)
Развитие диафрагмы
Развитие всех групп мышц - выполнение дифференцированных движений
2 Онтогенез мышечной системы: источники и сроки развития .
Производные миотома: из дорзального участка развиваются мышцы спины
из вентрального - мышцы груди и живота
Мезенхима - мышцы конечностей
I висцеральная дуга (ВД) - жевательные мышцы
II ВД - мимические мышцы
III и IV ВД - мышцы мягкого неба, глотки, гортани, верхней части пищевода
V ВД - грудино-ключично-сосцевидная и трапециевидная мышцы
Из затылочных миотомов - мышцы языка
Из предушных миотомов - мышцы глазного яблока
Мышцы развиваются из мезодермы . На туловище они возникают из первичной сегментированной мезодермы – сомитов : 3-5 затылочных, 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых, 4-5 копчиковых.
Каждый сомит подразделяется на склеротом, дерматом и миотом – из него и развиваются мышцы туловища. Сомиты появляются рано, когда длина зародыша составляет 10-15 мм.
Из дорсальных частей миотомов возникают глубокие, собственные (аутохтонные) мышцы спины, из вентральных – глубокие мышцы груди и живота. Они закладываются, развиваются и остаются в пределах туловища – поэтому называются аутохтонными (местными, туземными ). Очень рано миотомы связываются с нервной системой и каждому мышечному сегменту соответствует нервный сегмент. Каждый нерв следует за развивающейся мышцей, врастает в нее и, пока она не дифференцировалась, подчиняет своему влиянию.
В процессе развития часть скелетных мышц перемещается с туловища и шеи на конечности – трункофугальные мышцы: трапециевидная, грудино-ключично-сосцевидная, ромбовидные, поднимающая лопатку и др. Часть мышц, наоборот, направляется с конечностей на туловище – трункопетальные мышцы: широчайшая мышца спины, большая и малая грудные, большая поясничная.
Мышцы головы мимические и жевательные, над- и подъязычные мышцы шеи развиваются из несегментированной вентральной мезодермы, входящей в состав висцеральных (жаберных) дуг. Они называются висцеральными и, например, жевательные мышцы развиваются на основе первой висцеральной дуги, а мимические – второй. Однако мышцы глазного яблока и языка развиваются из затылочных миотомов сегментированной мезодермы. Глубокие передние и задние мышцы шеи также возникают из затылочных шейных миотомов, а поверхностная и средняя группа мышц в передней области шеи развиваются на основе несегментированной мезодермы висцеральных дуг.
3 Мышца: определение, строение .
Мускул (мышца) – орган, построенный из мышечных волокон (клеток), каждое из них обладает соединительно-тканной оболочкой – эндомизием . В пучки мышечные волокна объединяет другая фиброзная оболочка – перимизий , а весь мускул заключается в общую фиброзную оболочку, образованную фасцией – эпимизий . Между пучками проходят сосуды и нервы, снабжающие мышечные волокна.
На макроуровне скелетная мышца имеет:
· брюшко (вентер) – мясистая часть органа, занимающая его средину;
· сухожилие (тендо), относящееся к дистальному концу, оно может быть в виде апоневроза, сухожильных перемычек, длинных пучков продольных фиброзных волокон;
· головку , составляющую проксимальную часть;
· сухожилие и головка крепятся на противоположных концах костей.
Проксимальное сухожилие или головка мышцы - начало мышцы на кости находится ближе к срединной оси тела – это фиксированная точка (punctum fixum) (как правило совпадает с началом мышцы). Дистальное сухожилие, «хвост» - конец мышцы лежит на кости дистальнее и, являясь местом прикрепления, называется подвижной точкой (punctom mobile). При сокращении мышцы точки сближаются, а при изменении положения тела могут меняться местами.
Сухожилия разные по форме: тонкие длинные сухожилия имеют мышцы конечностей; мышцы, участвующие в формировании стенок брюшной полости, имеют широкое плоское сухожилие, расположенное меж двумя брюшками - сухожильное растяжение или апоневроз.
4 Классификация мышц по происхождению, строению, форме и функции .
А. Везалий – анатом эпохи Возрождения - обозначал мышцы цифрами, но сейчас они классифицируются по другим принципам.
По происхождению:
- из дорсальных частей миотомов возникают глубокие, собственные (аутохтонные) мышцы спины
- из вентральных – глубокие мышцы груди и живота, они закладываются, развиваются и остаются в пределах туловища – поэтому называются аутохтонными (местными, туземными ).
По функции различают:
· мышцы-антагонисты , как то: сгибатели и разгибатели, приводящие и отводящие, супинаторы и пронаторы – такие мышцы действуют в противоположных направлениях;
· мышцы-синергисты – действуют по одному направлению, усиливая друг друга; при ряде движений в качестве синергистов выступают и мышцы-антагонисты, например, при выполнении круговых движений;
· главные и вспомогательные мышцы.
По расположению:
· наружные и внутренние
· поверхностные и глубокие
· медиальные и латеральные
По форме и строению:
· веретенообразные мышцы (musculi fusiformes) – явл. длинными рычагами (двуглавая мышца плеча)
· широкие мышцы – участвуют в образовании стенок туловища (прямая мышца живота)
· одно-, дву- и многоперистые мышцы – в зависимости от того, по одну сторону от сухожилия или на две и более сторон ложатся мышечные пучки, например, многоперистая дельтовидная мышца.
· мышцы, форма которых соответствует определенной геометрической фигуре, например, ромбовидные большая и малая, трапециевидная, квадратная, круговая, прямая, тонкая;
· мышцы, имеющие несколько головок или брюшек: двух-, трех-, четырехглавые мышцы конечностей; двубрюшная на шее;
· мышцы, в названиях которых отражено направление волокон: поперечная, продольная, косая;
· мышцы, в названиях которых отражена функция: разгибатель, сгибатель, приводящая, отводящая, поднимающая, опускающая, сжимающая и т. д.;
· мышцы, большие по площади и длине: широкие и широчайшие, большие и малые, длинные и короткие;
· одно-, дву- и многосуставные мышцы в зависимости от того, на сколько суставов действуют мышцы, есть мышцы которые вообще не действуют на сустав.
Так же различают:
- гладкие мышцы (непроизвольная мускулатура) – развиваются из висцерального листка сплахнотома, располагаются в стенке внутренних органов, сокращаются самопроизвольно, иннервируются вегетативной нервной системой;
- поперечно-полосатые мышцы – скелетные (произвольная муск., разв. из миотомов – обр. скелет. мускулатуру, инерв.- сомат.нерв.сист) и сердечная (непроизвольная мускулатура, имеет попер-полос строен., но сост. из отдел. клеток – кардиомиоцитов, инерв. – вегет.нерв.сист.)
Скелетные мышцы выполняют преодолевающую, уступающую работу , что обеспечивает мышечную динамику тела, удерживающую – миостатическую работу.
АНАТОМИЯ ДОМАШНИХ ЖИВОТНЫХ
ПЛОСКОСТИ ТЕЛА И ТЕРМИНЫ ДЛЯ ОБОЗНАЧЕНИЯ РАСПОЛОЖЕНИЯ ОРГАНА
Для определения расположения органов и частей тело животного расчленяют тремя воображаемыми взаимоперпендикулярными плоскостями – сагиттальной, сегментальной и фронтальной (рис.1).
Срединная сагиттальная (медианная) плоскость проводится вертикально вдоль середины тела животного от рта до кончика хвоста и рассекающая его на две симметричные половины. Направление в теле животного к срединной плоскости называется медиальным, а от нее – латеральным (lateralis - боковой).
Рис.1. Плоскости и направления в теле животного
Плоскости:
I – сегментальная;
II - сагиттальная;
III – фронтальная.
Направления:
1 – краниальное;
2 – каудальное;
3 – дорсальное;
4 – вентральное;
5 – медиальное;
6 – латеральное;
7 – ростральное (оральное);
8 – аборальное;
9 – проксимальное;
10 – дистальвое;
11 – дорсальное
(спинковое, тыльное);
12 – пальмарное;
13 - плантарное.
Сегментальную плоскость проводят вертикально поперек тела животного. Направление от нее в сторону головы называется краниальным (cranium - череп), в сторону хвоста – каудальным (cauda - хвост). На голове, где все краниальное, различают направление к носу – назальное или хоботку – ростральное и противоположное ему-каудальное.
Фронтальную плоскость (frons - лоб) проводят горизонтально вдоль тела животного (при горизонтально вытянутой голове), т. е. параллельно лбу. Направление в данной плоскости в сторону спины называется дорсальным (dorsum - спина), к животу – вентральным (venter - живот).
Для определения положения участков конечностей существуют термины проксимальный (proximus - ближайший) – более близкое положение к осевой части тела и дистальный {distalus - удаленный) – более отдаленное положение от осевой части тела. Для обозначения передней поверхности конечностей приняты термины краниальный или дорсальный (для лапы), а для задней поверхности – каудальный, а также пальмарный или волярный (palma, vola - ладонь) – для кисти и плантарный (planta - стопа) – для стопы.
ОТДЕЛЫ И ОБЛАСТИ ТЕЛА ЖИВОТНОГО И ИХ КОСТНАЯ ОСНОВА
 |
Тело животных делят на осевую часть и конечности. Начиная с амфибий, у животных осевую часть тела делят на голову, шею, туловище и хвост. Шея, туловище и хвост составляют ствол тела. Каждая из частей тела разделена на отделы и области (рис.2). Основой их в большинстве случаев являются кости скелета, имеющие те же названия, что и области.
Рис. 2 Области тела крупного рогатого скота
1 - лобная; 2 - затылочная; 3 - теменная; 4 - височная; 5 - околоушная; 6 - ушной раковины; 7 - носовая; 8 - области верхней и нижней губ; 9 - подбородочная; 10 - щечная; 11 - межчелюстная; 12 - подглазничная; 13 - скуловая; 14 - область глаза; 15 - большой жевательной мышцы; 16 - верхняя шейная; 17 – боковая шейная; 18 - нижняя шейная; 19 - холки; 20 - спины; 21 - реберная; 22 - предгрудинная; 23 - грудинная: 24 - поясничная: 25 - подреберья; 26 - мечевидного хряща; 27 - околопоясничyая (голодная) ямка; 28 - боковая область; 29 - паховая; 30 - пупочная; 31 - лонная; 32 - маклок; 33 – крестцовая; 34 - ягодичная; 35 - корень хвоста; 36 - седалищная область; 37 - лопатка; 38 - плечо; 39 - предплечье; 40 - кисть; 41 - запястье; 42 - пясть; 43 - пальцы; 44 - бедро; 45 - голень; 46 - стопа; 47 - заплюсна; 48 - плюсна.
Голова (лат. caput, греч. cephale) делится на череп (мозговой отдел) и лицо (лицевой отдел). Череп (cranium) представлен областями: затылочной (затылок), теменной (темя), лобной (лоб) с областью рога у рогатого скота, височной (висок) и околоушной (ухо) с областью ушной раковины. На лице (facies) различают области: глазничную (глаза) с областями верхнего и нижнего век, подглазничную, скуловую с областью большой жевательной мышцы (у лошади – ганаши), межчелюстную, подбородочную, носовую (нос) с областью ноздрей, ротовую (рот), в состав которой входят области верхней и нижней губ и щеки. Над верхней губой (в области ноздрей) находится носовое зеркальце, у крупных жвачных оно распространяется на область верхней губы и становится носогубным.
Шея
Шея (cervix, collum) простирается от затылочной области до лопатки и делится на области: верхнюю шейную, лежащую над телами шейных позвонков; боковую шейную (область плечеголовной мышцы), идущую вдоль тел позвонков; нижнюю шейную, вдоль которой тянется яремный желоб, а также гортанную и трахейную (на вентральной ее стороне). У копытных шея сравнительно длинная в связи с необходимостью питания подножным кормом. Самая длинная шея у быстроаллюрных лошадей. Самая короткая – у свиньи.
Туловище
Туловище (truncus) состоит из грудного, брюшного и тазового отделов.
Грудной отдел включает области холки, спины, боковые реберные, предгрудинную и грудинную. Он прочный и подвижный. В каудальном направлении прочность уменьшается, а подвижность нарастает благодаря особенностям их соединения. Костной основой холки и спины являются грудные позвонки. В области холки у них наиболее высокие остистые отростки. Чем выше и длиннее холка, тем больше площадь прикрепления мышц позвоночника и пояса грудной конечности, тем размашистее и более упруги движения. Между длиной холки и спины существует обратная зависимость. Самая длинная холка и короткая спина у лошади, у свиньи – наоборот.
Брюшной отдел включает поясницу (lumbus), живот (abdomen), или брюхо (venter), поэтому еще называется пояснично-брюшным отделом. Поясница – продолжение спины до крестцовой области. Ее основой являются поясничные позвонки. Живот имеет мягкие стенки и делится на ряд областей: правого и левого подреберий, мечевидного хряща; парную боковую (подвздошную) с голодной ямкой, примыкающую снизу к пояснице, спереди – к последнему ребру, а сзади – переходит в паховую область; пупочную, лежащую снизу живота позади области мечевидного хряща и впереди от лонной области. На вентральной поверхности областей мечевидного хряща, пупочной и лонной у самок расположены молочные железы. У лошади наиболее короткая поясница и менее обширный брюшной отдел. У свиньи и рогатого скота поясница более длинная. Наиболее объемистый брюшной отдел у жвачных.
Тазовый отдел (pelvis) делится на области: крестцовую, ягодичную, включая маклок, седалищную и промежностную с примыкающей мошоночной областью. В хвосте (cauda) различают корень, тело и кончик. Области крестцовая, две ягодичных и корень хвоста у лошади образуют круп.
Конечности (membra) подразделяются на грудные (передние) и тазовые (задние). Состоят они из поясов, которыми соединяются со стволовой частью тела, и свободных конечностей. Свободные конечности делятся на основной поддерживающий столб и лапу. Грудная конечность состоит из плечевого пояса, плеча, предплечья и кисти.
Области плечевого пояса и плеча примыкают к боковой грудной области. Костной основой плечевого пояса у копытных является лопатка, поэтому его часто называют областью лопатки. Плечо (brachium) расположено ниже плечевого пояса, имеет форму треугольника. Костной основой является плечевая кость. Предплечье (antebrachium) находится вне кожного туловищного мешка. Его костная основа – лучевая и локтевая кости. Кисть (manus) состоит из запястья (carpus), пясти (metacarpus) и пальцев (digiti). У животных разных видов их бывает от 1 до 5. Каждый палец (кроме первого) состоит из трех фаланг: проксималыной, средней и дистальной (которые у копытных называются соответственно путовой, у лошади – бабка), венечной и копытцевой (у лошади – копытной).
Тазовая конечность состоит из тазового пояса, бедра, голени и стопы.
Область тазового пояса (таз) входит в состав осевой части тела в качестве ягодичной области. Костная основа – тазовая или безымянная кости. Область бедра (femur) расположена под тазом. Костная основа – бедренная кость. Область голени (crus) находится вне кожного туловищного мешка. Костная основа - большая и малая берцовые кости. Стопа (pes) состоит из заплюсны (tarsus), плюсны (metatarsus) и пальцев (digiti). Количество их, строение и названия у копытных такие же, как на кисти.
СОМАТИЧЕСКИЕ СИСТЕМЫ
Кожный покров, скелетная мускулатура и скелет, формируя собственно тело – сому животного,-объединяются в группу соматических систем организма.
Аппарат движения образован двумя системами: костной и мышечной. Кости, объединенные в скелет, представляют собой пассивную часть аппарата движения, являясь рычагами, на которые действуют прикрепленные к ним мышцы. Мышцы действуют только на кости, подвижно соединенные с помощью связок. Мышечная система – активная часть аппарата движения. Он обеспечивает движение организма, его перемещение в пространстве, поиск, захват и пережевывание пищи, нападение и защиту, дыхание, движения глаз, ушей и др. На его долю приходится от 40 до 60% массы организма. Он определяет форму тела животного (экстерьер), пропорции, обусловливая типовые особенности конституции, что имеет огромное практическое значение в зоотехнии, т. к. с особенностями экстерьера, типом конституции связаны выносливость, приспособляемость, способность к откорму, скороспелость, половая активность, жизнестойкость, и другие качества животных.
СКЕЛЕТ, СОЕДИНЕНИЕ КОСТЕЙ СКЕЛЕТА (ОСТЕОЛОГИЯ)
Общая характеристика и значение скелета.
Скелет (греч. skeleton - иссохший, мумия) образован костями и хрящами, соединенными между собой соединительной, хрящевой или костной тканями. Скелет млекопитающих называется внутренним, т. к. он расположен под кожей и покрыт слоем мышц. Он является твердой основой тела и служит футляром для головного, спинного и костного мозга, для сердца, легких и других органов. Упругость и рессорные свойства скелета обеспечивают плавность движений, предохраняют мягкие органы от толчков и сотрясений. Скелет участвует в минеральном обмене. В нем содержатся большие запасы солей кальция, фосфора и др. веществ. Скелет – наиболее точный показатель степени развития и возраста животного. Многие прощупываемые кости являются постоянными ориентирами при проведении зоотехнических измерений животного.
ДЕЛЕНИЕ СКЕЛЕТА
Скелет делится на осевой и скелет конечностей (периферический) (рис.3).
Осевой скелет включает скелет головы, шеи, туловища и хвоста. Скелет туловища состоит из скелета грудной клетки, поясницы и крестца. Периферический скелет образован костями поясов и свободных конечностей. Количество костей у животных разных видов, пород и даже особей неодинаково. Масса скелета составляет у взрослого животного от 6% (свиньи) до 12-15% (лошадь, бык). У новорожденного телят – до 20%, а у поросят – до 30%. от массы тела. У новорожденных более развит периферический скелет. На его долю приходится 60-65% массы всего скелета, а на долю осевого 35-40%. После рождения активнее растет, особенно в молочный период, осевой скелет и у 8-10-месячного теленка отношения этих отделов скелета выравниваются, а затем осевой начинает преобладать: в 18 месяцев у крупного рогатого скота он составляет 53-55%. У свиньи масса осевого и периферического скелета примерно одинакова.

 |
 Рис.3 Скелет коровы (А), свиньи (Б),
Рис.3 Скелет коровы (А), свиньи (Б),
лошади (В)
Осевой скелет: 1- кости мозгового отдела (черепа): 3- кости лицевого отдела (лица); а- шейные позвонки; 4 - грудные позвонки; 5 - ребра; 6 - грудина; 7 - поясничные позвонки: 8 - крестцовая кость: 9 - хостовые позвонки (3,4,7,8,9 - позвоночник). Скелет конечностей; 10 - лопатка; 11 - плчевая кость; 12 - кости предплечья (лучевая и локтевая); 13 - кости запястья; 14 - кости пясти; 15 - кости пальцев (IS-15 – кости кисти); 16 - тазовая кость; П - бедренная кость: IS - коленная чашка; IS - кости голени (большая и малая берцовые); 30 - кости заплюсны: 31 - кости плюсны; 32 - кости пальцев (20-22 - кости стопы).
Форма и строение костей
Кость (лат. os) – орган костной системы. Как всякий орган, она имеет определенную форму и состоит из нескольких видов тканей. Форма костей определяется особенностями ее функционирования и положением в скелете. Различают длинные, короткие, плоские и смешанные кости.
Длинные кости бывают трубчатыми (многие кости конечностей) и дугообразными (ребра). Длина тех и других больше ширины и толщины. Длинные трубчатые кости напоминают по форме цилиндр с утолщенными концами. Средняя, более узкая часть кости называется телом – диафиз (греч. diaphysis), расширенные концы – эпифизы (epiphysis). Эти кости играют основную роль в статике и динамике, в кроветворной функции (содержат красный костный мозг).
Короткие кости обычно небольшой величины, их высота, ширина и толщина близки по размеру. Они часто выполняют рессорную функцию.
Плоские кости имеют большую поверхность (ширину и длину) при малой толщине (высоте). Обычно они служат стенками полостей, защищая помещенные в них органы (черепномозговая коробка) или это обширное поле для прикрепления мышц (лопатка).
Смешанные кости имеют сложную форму. Эти кости, как правило, непарные и размещаются по оси тела. (затылочная, клиновидная кости, позвонки). Парные смешанные кости несимметричны, например височная кость.
Строение кости
Основная ткань, образующая кость – пластинчатая костная. В состав кости входят также ретикулярная, рыхлая и плотная соединительные ткани, гиалиновый хрящ, кровь и эндотелий сосудов, нервные элементы.
Снаружи кость одета надкостницей, или периостом, за исключением места расположения суставного хряща. Наружный слой надкостницы фиброзный, образован соединительной тканью с большим количеством коллагеновых волокон; определяет ее прочность. Внутренний слой содержит недифференцированные клетки, которые могут преобразовываться в остеобласты и являются источником роста кости. Через надкостницу в кость проникают сосуды и нервы. Надкостница во многом определяет жизнеспособность кости. Кость, очищенная от надкостницы, погибает.
Под надкостницей залегает слой кости, образованный плотно уложенными костными пластинками. Это компактное вещество кости. В трубчатых костях в нем различают несколько зон. К надкостнице примыкает зона наружных генеральных пластинок толщиной 100-200 мкм. Она придает кости большую твердость. Затем следует наиболее широкая и важная в структурном отношении зона остеонов. Чем толще слой остеонов, тем лучше рессорные свойства кости. В этом слое между остеонами залегают вставочные пластинки – остатки старых разрушенных остеонов. У копытных в нем часто встречаются циркулярно-параллельные структуры, устойчивые к сопротивлению на изгиб. Не случайно они широко распространены в длинных трубчатых костях копытных, испытывающих большое давление. Толщина внутреннего слоя компактного вещества 200-300 мкм, образован он внутренними генеральными пластинками или же переходит в губчатое вещество кости.
Губчатое вещество представлено костными пластинками, которые не плотно примыкают друг к другу, а формируют сеть из костных перекладин (трабекул), в ячеях которой располагается красный костный мозг. Губчатое вещество особенно развито в эпифизах. Его перекладины располагаются не беспорядочно, а строго следуют линиям действующих сил (сжатия и растяжения).
В середине диафиза трубчатой кости имеется костная полость . Она образовалась в результате резорбции костной ткани остеокластами во время развития кости и заполнена желтым (жировым) костным мозгом.
Кость богата сосудами, которые образуют сеть в ее надкостнице, пронизывают всю толщину компактного вещества, находясь в центре каждого остеона, и разветвляются в костном мозге. В кости кроме сосудов остеонов имеются т. н. питательные сосуды (фолькмановы), прободащие кость перпендикулярно ее длине. Вокруг них не образуется концентрических костных пластинок. Особенно много таких сосудов около эпифизов. Нервы в кость проникают от надкостницы через те же отверстия, что и сосуды. Поверхность кости покрыта гиалиновым хрящом без надхрящницы. Толщина его 1-6 мм и прямо пропорциональна нагрузке на сустав.
Строение коротких, сложных и плоских костей такое же, как и трубчатых, с той лишь разницей, что у них обычно нет костных полостей. Исключение составляют некоторые плоские кости головы, в которых между пластинками компактного вещества имеются обширные пространства, заполненные воздухом – синусы или пазухи.
ФИЛОГЕНЕЗ СКЕЛЕТА
Развитие опорной системы в филогенезе животных шло двумя путями: образования наружного и внутреннего скелета. Наружный скелет закладывается в покровах тела (членистоногие). Внутренний скелет развивается под кожей и обычно прикрыт мышцами. О развитии внутреннего скелета можно говорить со времени появления хордовых. У примитивных хордовых (ланцетник) – хорда представляет собой опорную систему. С усложнением организации животных соединительнотканный скелет замещается хрящевым, а затем костным.
Филогенез стволового скелета
В филогенезе позвоночных раньше других элементов появляются позвонки. С усложнением организации, увеличением активности и разнообразия движений вокруг хорды развиваются не только дуги, но и тела позвонков. У хрящевых рыб скелет образован хрящом, иногда обызвествленным. Кроме верхних дуг под хордой у них развиваются нижние дуги. Концы верхних дуг каждого сегмента, сливаясь, образуют остистый отросток. Появляются тела позвонков. Хорда теряет значение опорного стержня. У костистых рыб происходит замещение хрящевого скелета костным. Появляются суставные отростки, которыми позвонки сочленяются друг с другом, чем обеспечивается прочность скелета при сохранении его подвижности. Осевой скелет разделяется на головной, туловищный с ребрами, охватывающими полость тела с органами, и сильно развитый хвостовой отдел – локомоторный.
Переход к наземному образу жизни приводит к развитию одних частей скелета и редукции других. Скелет туловища дифференцируется на шейный, грудной (спинной), поясничный и крестцовый отделы, частично редуцируется скелет хвоста, т. к. основная нагрузка при движении по земле падает на конечности. В грудном отделе в тесной связи с ребрами развивается грудина, формируется грудная клетка. У земноводных шейный и крестцовый отделы позвоночника имеют только по одному позвонку, поясничный отдел отсутствует. Ребра очень короткие, у многих срастаются с поперечными отростками позвонков. У рептилий шейный отдел удлиняется до восьми позвонков и приобретает большую подвижность. В грудном отделе 1-5 пар ребер соединены с грудиной – образуется грудная клетка. Поясничный отдел длинный, имеет ребра, величина которых уменьшается в каудальном направлении. Крестцовый отдел образован двумя позвонками, хвостовой отдел длинный, хорошо развит.
У млекопитающих, независимо от образа жизни, постоянное число шейных позвонков (7). Относительно постоянно число позвонков и в остальных отделах: 12-19 грудных, 5-7 поясничных, 3-9 крестцовых. Хвостовых позвонков насчитывают от 3 до 46. Позвонки, за исключением первых двух, соединены при помощи хрящевых дисков (менисков), связок и суставных отростков.
Поверхности тел шейных позвонков часто имеют выпукло-вогнутую форму – опистоцельные. В остальных отделах позвонки обычно плоские-платицельные. Ребра сохраняются только в грудном отделе. В пояснице они редуцируются и срастаются с поперечными отростками позвонков. В крестцовом отделе срастаются и позвонки, образуя крестцовую кость. Хвостовой отдел облегчен, его позвонки сильно редуцируются.
Филогенез скелета головы
Скелет головного конца тела развивается вокруг нервной трубки – осевой (мозговой) скелет головы и вокруг головной кишки – висцеральный. Осевой скелет головы представлен хрящевыми пластинками, окружающими нервную трубку снизу и с боков, крыша черепа перепончатая. Висцеральный скелет головы состоит из хрящевых жаберных дуг, связанных с аппаратом дыхания и пищеварения; челюстей нет. Развитие скелета головы шло путем объединения мозгового и висцерального скелетов и усложнения их строения в связи с развитием головного мозга, органов чувств (обоняния, зрения, слуха). Мозговой череп хрящевых рыб представляет собой сплошную хрящевую коробку, окружающую головной мозг. Висцеральный скелет образован хрящевыми жаберными дугами. Черепная коробка у костистых рыб сложного строения. Первичные кости формируют затылочную область, часть основания черепа, обонятельную и слуховую капсулы и стенку глазницы. Покровные кости покрывают первичную черепную коробку сверху, снизу и с боков. Висцеральный скелет представляет собой очень сложную систему рычагов, участвующих в хватательных, глотательных и дыхательных движениях. С черепной коробкой висцеральный скелет сочленяется при помощи подвеска (hyomandibulare), в результате чего образуется единый скелет головы.
С выходом на сушу, с резкой сменой среды обитания и образа жизни животных происходят значительные изменения в скелете головы: череп подвижно причленяется к шейному отделу; уменьшается количество костей черепа за счет их сращения; увеличивается его прочность. Смена типа дыхания (с жаберного на легочное) приводит к редукции жаберного аппарата и преобразованию его элементов в подъязычную и слуховые кости. Челюстной аппарат срастается с основанием черепа. В ряду наземных животных прослеживается постепенное усложнение. В черепе земноводных много хрящей, слуховая кость одна. Для черепа млекопитающих характерно уменьшение количества костей за счет их сращения (например, затылочная кость образуется путем сращения 4, а каменистая – 5 костей), в стирании граней между первичными и покровными (вторичными) костями, в мощном развитии обонятельной области и сложном звукопроводящем аппарате, в крупных размерах черепной коробки и др.
Филогенез скелета конечностей
Гипотеза о происхождении конечностей наземных животных на базе парных плавников рыб сейчас имеет широкое признание. Парные плавники в типе хордовых впервые появились у рыб. Костной основой парных плавников рыб служит система хрящевых и костных элементов. Тазовый пояс у рыб развит в меньшей степени. С выходом на сушу, на базе парных плавников развивается скелет конечностей, расчлененный на отделы, типичные для пятипалой конечности. Пояса конечностей состоят из 3 пар костей и укрепляются связью с осевым скелетом: плечевой пояс – с грудиной, тазовый с крестцом. Плечевой пояс состоит из коракоида, лопатки и ключицы, тазовый – из подвздошной, лонной и седалищной костей. Скелет свободных конечностей расчленен на 3 отдела: у передней конечности –это кости плеча, предплечья и кисти, у задней – бедра, голени и стопы.
Дальнейшие преобразования связаны с характером передвижения, его скоростью и маневренностью. У земноводных пояс грудных конечностей, причленяясь к грудине, не имеет жесткой связи с осевым скелетом. В поясе тазовых конечностей развита его вентральная часть. У рептилий в скелете поясов в равной степени развиты дорсальная и вентральная части.
Плечевой пояс млекопитающих редуцирован и состоит из двух или даже одной кости. У животных с развитыми отводящими движениями грудной конечности (например, кротов, летучих мышей, обезьян) развиты лопатка и ключица, а у животных с однообразными движениями (например, у копытных) – только лопатка. Тазовый пояс млекопитающих упрочен тем, что лонные и седалищные кости соединяются вентрально с соименными костями. Скелет свободных конечностей млекопитающих организован так, что тело животного поднято над землей. Приспособление же к различным видам передвижения (бег, лазанье, прыжки, полет, плавание) привело к сильной специализации конечностей у разных групп млекопитающих, что выражается в основном в изменении длины и угла наклона отдельных звеньев конечностей, формы суставных поверхностей, срастании костей и редукции пальцев.
Изменение в строении конечностей в филогенезе в связи с повышением специализации – приспособленностью к определенному виду движения подробнее всего изучено в ряду лошадей (). Предполагаемый предок лошади, совмещая в себе черты копытных и хищных, был размером с лисицу и имел пятипалые конечности с когтями, по форме приближающимися к копытцам. От разнообразных мягких движений по рыхлой земле с высокой растительностью (лес) до широких размашистых быстрых движений по сухим открытым пространствам (степь) происходило удлинение основного поддерживающего столба конечностей из-за раскрытия (увеличения) углов между ее звеньями. Лапа приподнималась, животное переходило от стопо - к пальцехождению. При этом наблюдалась постепенная редукция нефункционирующих пальцев. При переходе от пальце - к фаланго (копыто-) хождению в состав основного поддерживающего столба входит уже вся лапа, а редукция пальцев достигает максимума. У лошади полноразвитым сохраняется на конечности лишь III палец. У рогатого скота развитыми оказываются два пальца-III и IV.
Онтогенез скелета
В процессе индивидуального развития особи скелет проходит те же 3 стадии развития и в той же последовательности, что и в филогенезе: соединительнотканного, хрящевого и костного скелета.
Хорда как один из первых осевых органов закладывается в зародышевый период внутриутробного развития в результате дифференцировки энтодермы и мезодермы в период гаструляции. Вскоре вокруг нее образуется сегментированная мезодерма – сомиты, внутренняя часть которых – склеротомы, прилежащие к хорде, являются скелетогенными зачатками.
Соединительнотканная стадия. В области склеротомов идет активное размножение клеток, которые принимают вид мезенхимных, разрастаются вокруг хорды и превращаются в ее соединительнотканный футляр и в миосепты – соединительнотканные тяжи. Соединительнотканный скелет у млекопитающих существует очень короткое время, так как параллельно процессу обрастания хорды в перепончатом скелете происходит размножение мезенхимных клеток, особенно вокруг миосепт, и дифференцировка их в хрящевые.
Хрящевая стадия. Дифференцировка мезенхимных клеток в хрящевые начинается с шейной области. Первыми закладываются хрящевые дуги позвонков, которые образуются между хордой и спинным мозгом, обрастают спинной мозг сбоку и сверху, формируя его футляр. Смыкаясь между собой попарно над спинным мозгом, дуги формируют остистый отросток. В это же время из сгущений мезенхимных клеток, размножающихся в футляре хорды, развиваются хрящевые тела позвонков, а в миосептах – зачатки ребер и грудины. Замещениея соединительной ткани хрящом начинается у свиньи и овцы на 5-й, у лошади и крупного рогатого скота – на 6-й неделе эмбрионального развития. Затем в такой же последовательности, в какой шло образование хрящевого скелета, идет его окостенение.
В хрящевой закладке (модели) кости сосудов нет. С развитием кровеносной системы зародыша идет образование сосудов вокруг и внутри надхрящницы, в результате чего ее клетки начинают дифференцироваться не в хондробласты, а в остеобласты, т. е. она становится надкостницей – периостом. Остеобласты продуцируют межклеточное вещество и откладывают его поверх хрящевого зачатка кости. Формируется костная манжетка. Костная манжетка построена из грубоволокнистой костной ткани. Процесс образования и роста манжетки вокруг хрящевого зачатка называется окостенением.
Костная манжетка затрудняет питание хряща и начинает разрушаться. Первые очаги обызвествления и деструкции хряща обнаруживают в центре (диафизе) хрящевого зачатка. В очаг разрушающегося хряща от надкостницы проникают сосуды вместе с недифференцированными клетками. Здесь они размножаются и превращаются в костные клетки – возникает первый очаг (центр) окостенения. У каждой кости обычно несколько очагов окостенения (в позвонках копытных их насчитывают 5-6, в ребрах – 1-3).
В очаге окостенения остеокласты разрушают обызвествленный хрящ, бразуя лакуны и туннели, шириной 50-800 мкм. Остеобласты вырабатывают межклеточное вещество, которое откладывается по стенкам лакун и туннелей. Мезенхима, проникающая вместе с капиллярами, дает начало следующей генерации остеобластов, которые, откладывая межклеточное вещество в сторону стенок туннелей, замуровывают предыдущие генерации остеобластов – развиваются костные пластинки. Поскольку лакуны и туннели образуют сеть, костная ткань, выстилающая их, повторяет их форму и в целом напоминает губку, состоящую из переплетающихся костных тяжей, перекладин или трабекул. Из них формируется губчатое вещество кости. Образование кости внутри хрящевого зачатка на месте разрушенного хряща называется эндохондральным (энхондральным) окостенением.
Часть недифференцированных клеток, проникающих вместе с капиллярами в туннели и лакуны, превращается в клетки костного мозга, который заполняет пространства между костными трабекулами губчатого вещества.
Процесс энхондрального окостенения, начавшись в области диафиза, распространяется к концам зачатка – эпифизам. Параллельно с этим утолщается и нарастает костная манжетка. В таких условиях хрящевая ткань может расти только в продольном направлении. При этом хондробласты, размножаясь, выстраиваются друг над другом в виде клеточных колонок (монетных столбиков).
Закладка хрящевый моделей и их окостенение происходят быстро в тех участках тела, где потребность в опоре появляется очень рано. По срокам закладки и темпам дифференцировки костного скелета млекопитающих можно разделить на несколько групп. Копытные относятся к группе, у которой закладка и формирование очагов окостенения почти заканчиваются к моменту рождения, 90% кости образовано костной тканью. После рождения продолжается лишь рост этих очагов. Новорожденные таких животных активны, сразу могут самостоятельно передвигаться, следовать за матерью и добывать себе корм.
Первичные очаги окостенения в предплодный период отмечают в скелете туловища. У рогатого скота вначале окостеневают ребра. Окостенение позвонков начинается с атланта и распространяется в каудальном направлении. Тела окостеневают прежде всего у средних грудных позвонков. Во второй половине эмбрионального развития активна образуются остеоны, выделяются слои наружных и внутренних генеральных пластинок. В постнатальном онтогенезе идет нарастание новых слоев костной ткани до завершения роста животного, а также перестройка уже существующих остеонов.
Зона клеточных колонок постоянно нарастает со стороны эпифизов благодаря дифференцировке хрящевых клеток от надхрящницы. Со стороны же диафиза отмечается постоянное разрушение хряща из-за нарушения его питания и изменения химизма ткани. До тех пор, пока эти процессы уравновешивают друг друга, кость растет в длину. Когда же скорость энхондрального окостенения становится больше, чем скорость роста метаэпифизарного хряща, он истончается и полностью исчезает. С этого времени прекращается линейный рост животного. В осевом скелете дольше всего сохраняются хрящи между эпифизами и телом позвонков, особенно в крестце.
В энхондральной кости рост кости в ширину начинается из диафиза и выражается в разрушении старых и образовании новых остеонов, в формировании костной полости. В перихондральной кости перестройка заключается в том, что грубоволокнистая костная ткань манжетки замещается пластинчатой костной тканью в виде остеонов, циркулярно -параллельных структур и генеральных пластинок, составляющих в совокупности компактное вещество кости. В процессе перестройки образуются вставочные пластинки. У крупного рогатого скота и свиньи осевой скелет начинает окостеневать в 3-4 года, а полностью процесс завершается в 5-7 лет, у лошади – в 4-5 лет, у овцы – в 3-4 года.
Развитие черепа
Начало осевому черепу дают 7-9 сомитов. Вокруг конечного участка хорды склеротомы этих сомитов образуют сплошную перепончатую пластинку без следов сегментации. Она распространяется вперед (прехордально) и охватывает снизу и с боков мозговые пузыри, слуховые и обонятельные капсулы и глазные бокалы. Замещение соединительнотканного осевого черепа хрящевым начинается около переднего конца хорды под основанием головного мозга. Здесь закладывается пара околохордовых (парохордалии) хрящей. Дальше в оральном направлении закладываются две хрящевые балки или трабекулы. Т. к. они лежат впереди хорды, этот отдел осевого черепа называется прехордальным. Трабекулы и парахордалии, разрастаясь, сливаются вместе, образую основную хрящевую пластинку. В оральной части вдоль основной хрящевой пластинки закладывается хрящевая носовая перегородка, по обе стороны от которой развиваются носовые раковины. Затем хрящи замещаются первичными, или примордиальными, костями. Первичными костями осевого черепа являются затылочная, клиновидная, каменистая и решетчатая, образующие дно, переднюю и заднюю стенки черепномозговой полости, а также носовая перегородка и раковины. Остальные кости вторичные, кожные, или покровные, т. к. возникают из мезенхимы, минуя хрящевую стадию. Это теменные, межтеменные, лобные, височные (чешуя), формирующие крышу и боковые стенки черепномозговой полости.
Параллельно развитию осевого черепа идет преобразование висцерального скелета головы. Большая часть зачатков висцеральных дуг подвергается полной редукции, а часть их материала идет на образование слуховых косточек, подъязычной кости и хрящей гортани. Основная же масса костей висцерального скелета – вторичные, покровные. Осевой и висцеральный скелет головы млекопитающих настолько тесно связывается друг с другом, что кости одного входят в состав другого. Поэтому череп млекопитающих делят на мозговой отдел (собственно череп), являющийся вместилищем мозга, и лицевой отдел (лицо), образующий стенки носовой и ротовой полостей. В плодный период определяется форма черепа, характерная для вида и породы. Роднички – неокостеневшие участки – закрываются плотной соединительной тканью или хрящом.
Развитие конечностей
Конечности у млекопитающих закладываются в виде выростов шейно-грудных и пояснично-крестцовых сомитов. У КРС это происходит на 3-й неделе. Сегментация их не выражена. Закладки имеют вид скоплений мезенхимы, которые быстро увеличиваются в длину, превращаясь в лопастевидные выросты. Сначала эти выросты подразделяются на два звена: закладку поясов и свободных конечностей, не расчлененных на отделы и кости. Затем из сгущений мезенхимы дифференцируются соединительнотканные и хрящевые закладки костей. В процессе дифференцировки скелет конечностей проходит те же три стадии, что и стволовой скелет, но с некоторым отставанием. Окостенение конечностей у плода теленка начинается на 8-9-й неделе и протекает аналогично со стволовым скелетом. Многие выросты костей – апофизы. имеют собственные очаги окостенения. В процессе окостенения в трубчатых костях образуется губчатое и компактное вещество. Перестройка из центра кости распространяется на ее периферию. При этом в области диафиза благодаря деятельности остеокластов почти полностью исчезает губчатое вещество, сохраняясь лишь в эпифизах. Увеличивается костная полость. Красный костный мозг в ней становится желтым.
Слои компактного вещества становятся заметны в течение первых месяцев жизни. Степень его развития зависит от вида животного. У копытных в нем хорошо развиты генеральные пластинки и циркулярно-параллельные структуры, у хищных преобладают остеоны. Это связано с различиями в функциональных нагрузках костей, особенно конечностей. У копытных они приспособлены к прямолинейному движению и удержанию массивного тела, у хищных – к более легкому телу и разнообразным движениям.
В конечностях очаги окостенения появляются в костях поясов, распространяясь затем в дистальном направлении. Окончательное окостенение (синостоз) прежде всего происходит в дистальных звеньях. Так, у крупного рогатого скота окостенение дистальных звеньев конечности (плюсны и пясти) завершается к 2-2,5 годам, к 3-3,5 годам окостеневают все кости свободной конечности, а кости тазового пояса – лишь к 7 годам.
Возрастные изменения скелета
В связи с разными сроками закладки, скорости роста и окостенения костей скелета на протяжении онтогенеза происходит изменение пропорций тела. В период эмбрионального развития кости растут с разной интенсивностью. У копытных в первую половину интенсивнее растет осевой скелет, во вторую – скелет конечностей. Так, у 2-месячных плодов телят осевой скелет составляет 77%, скелет конечностей – 23%, а к рождению – 39 и 61%. По данным, у мериноса от времени хрящевой закладки (1-месячный эмбрион) до рождения скелет тазовой конечности с поясом увеличивается в 200 раз, грудной конечности – в 181 раз, таз – в 74 раза, позвоночник – в 30 раз, череп – в 24 раза. После рождения на смену усиленному росту периферического скелета приходит линейный рост осевого скелета.
В постнатальном онтогенезе скелет растет с меньшей скоростью, чем мышцы и многие внутренние органы, поэтому его относительная масса снижается в 2 раза. В процессе роста и дифференцировки костей увеличивается их прочность, что связано с увеличением число остеонов на единицу площади. От рождения до взрослого состояния толщина компактного вещества увеличивается в 3-4 раза, содержание в нем минеральных солей – в 5 раз, предельная нагрузка – в 3-4 раза, достигая у овец 280, у коров 1000 кг на 1 см2. Окончательной прочности кости крупного рогатого скота достигают уже к 12-месячному возрасту.
Xем крупнее животное, тем меньше у него запас прочности костей. У самцов кости толще, чем у самок, но недокорм сказывается на них сильнее. Улучшенные породы овец и свиней имеют более короткие и широкие кости конечностей. У скороспелых животных кости толще, чем у позднеспелых. Кости коров молочного типа лучше снабжаются кровью, а у коров мясного и мясо-молочного типов больше площадь компактного вещества кости и толщина стенки, что обусловливает большую прочность еее при нагрузках. Прочность кости на изгиб обусловливает структура остеонов. У свиней породы ландрас, например, прочность кости на изгиб выше, чем у крупной белой и сибирской северной пород, в связи с тем, что у ландрас более плотное расположение остеонов.
Из всех внешних условий на развитие скелета наибольшее влияние оказывает кормление и моцион. Улучшение кормления в период интенсивного роста костей ускоряет, недокорм угнетает их темпы роста, особенно в ширину, но не нарушает общих закономерностей роста скелета. У животных пастбищного содержания компактное вещество кости плотнее, в нем преобладают пластинчатые структуры, трабекулы губчатого вещества толще, равномернее по ширине и направлены строго по действию сил сжатия - растяжения. При стойловом и клеточном содержании животных замедляется рост и внутренняя перестройка костей, снижается их плотность и прочность по сравнению с выгульным, напольным содержанием и с животными, подвергающимися дозированному принудительному движению.
Добавление к рациону молодняка макро - и микроэлементов способствует образованию костей с более толстым компактным веществом и трабекулами и меньшей костной полостью. При недостатке минеральных веществ происходит деминерализация скелета, размягчение и резорбция позвонков, начиная с хвостовых.
ЛЕКЦИЯ МИОЛОГИЯ ФИЛОГЕНЕЗ, ОНТОГЕНЕЗ И ФУНКЦИОНАЛЬНАЯ АНАТОМИЯ МЫШЕЧНОЙ СИСТЕМЫ Выполнили: Владимирова Я. Б. Кокорева Т. В.

 Мышцы или мускулы (от лат. musculus - мышка, маленькая мышь) - органы тела животных и человека, состоящие из упругой, эластичной мышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 86, 3 % из воды. В теле человека 640 мышц
Мышцы или мускулы (от лат. musculus - мышка, маленькая мышь) - органы тела животных и человека, состоящие из упругой, эластичной мышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 86, 3 % из воды. В теле человека 640 мышц
 Мотивация: - - - возможности совершаемого движения, объём движения; активные или пассивные движения запускаются в действие той или иной мышечной группой; воздействуя на мышечный аппарат, мы изменяем общее состояние; мышечный рельеф является ориентиром для топографии сосудов и нервов; пересадка мышц, то есть мышца может «переучиваться» .
Мотивация: - - - возможности совершаемого движения, объём движения; активные или пассивные движения запускаются в действие той или иной мышечной группой; воздействуя на мышечный аппарат, мы изменяем общее состояние; мышечный рельеф является ориентиром для топографии сосудов и нервов; пересадка мышц, то есть мышца может «переучиваться» .
 Развитие мышц Краниального происхождения - из головных миотомов (склеротомов) и мезенхимы жаберных дуг. Иннервируются ветвями черепномозговых нервов Спинального происхождения - из миотомов туловищного отдела зародыша: из вентральных миотомов иннервируются передними ветвями СМН; - из дорзальных миотомов иннервируются задними ветвями СМН - Аутохтонные мышцы – мыщцы, оставшиеся на месте своей первичной закладки. Трункофугальные мышцы – мыщцы, переместившиеся с туловища на конечности. Трункопетальные мышцы – мыщцы, переместившиеся с конечностей на туловище.
Развитие мышц Краниального происхождения - из головных миотомов (склеротомов) и мезенхимы жаберных дуг. Иннервируются ветвями черепномозговых нервов Спинального происхождения - из миотомов туловищного отдела зародыша: из вентральных миотомов иннервируются передними ветвями СМН; - из дорзальных миотомов иннервируются задними ветвями СМН - Аутохтонные мышцы – мыщцы, оставшиеся на месте своей первичной закладки. Трункофугальные мышцы – мыщцы, переместившиеся с туловища на конечности. Трункопетальные мышцы – мыщцы, переместившиеся с конечностей на туловище.


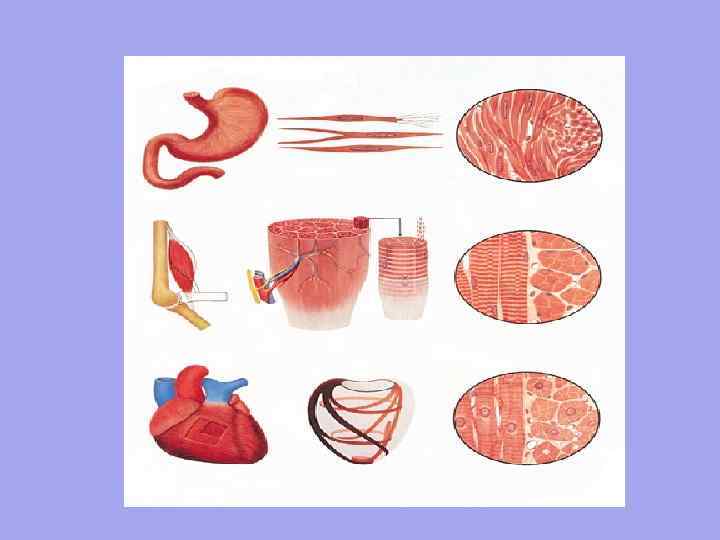
 Поперечно-полосатая Гладкая 1. Единицей организации является миоцит. Длина около 50 мкм. Ширина от 6 мкм. 2. Непроизвольное сокращение Контроль со стороны автономной нервной системы Движение волнообразное, работает медленно, так как нервное волокно подходит НЕ к каждой клетке Медленно вступают в действие, но сохраняются на длительном протяжении Не имеет точной пространственной ориентации клеток 3. 4. 5. 6. 1. 2. 3. 4. 5. 6. Сердечная Единицей организации является мышечное волокно- множество миобластов, плавающих в общей цитоплазме (саркоплазме). У них общая сарколемма. Длина около 40 -100 мм. Ширина от 7 мм. Произвольное сокращение Контроль со стороны соматической нервной системы Быстрое сокращение, быстрая реакция, так каждое мышечное волокно имеет нервно -мышечный синапс Быстро включаются, но имеет кратковременное действие Четкая ориентация мышечных волокон
Поперечно-полосатая Гладкая 1. Единицей организации является миоцит. Длина около 50 мкм. Ширина от 6 мкм. 2. Непроизвольное сокращение Контроль со стороны автономной нервной системы Движение волнообразное, работает медленно, так как нервное волокно подходит НЕ к каждой клетке Медленно вступают в действие, но сохраняются на длительном протяжении Не имеет точной пространственной ориентации клеток 3. 4. 5. 6. 1. 2. 3. 4. 5. 6. Сердечная Единицей организации является мышечное волокно- множество миобластов, плавающих в общей цитоплазме (саркоплазме). У них общая сарколемма. Длина около 40 -100 мм. Ширина от 7 мм. Произвольное сокращение Контроль со стороны соматической нервной системы Быстрое сокращение, быстрая реакция, так каждое мышечное волокно имеет нервно -мышечный синапс Быстро включаются, но имеет кратковременное действие Четкая ориентация мышечных волокон

 Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани- эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани- эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
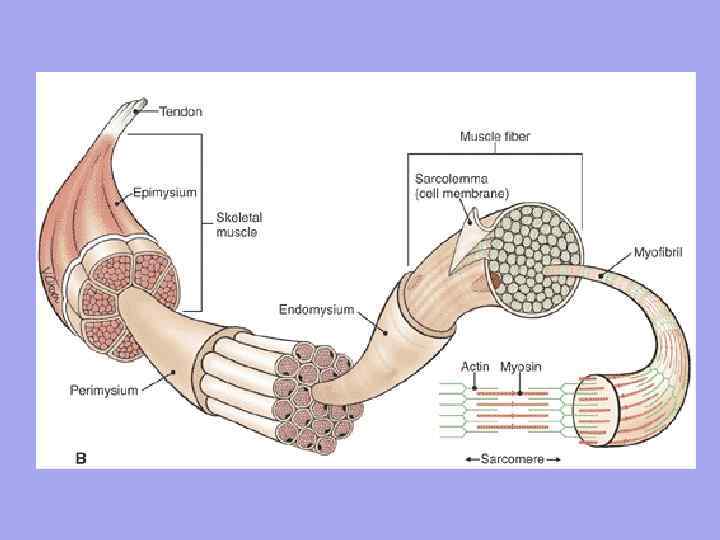
 В мышце как органе есть соединительная ткань Эндомизий – тонкая соединительная ткань, окружает каждое мышечное волокно и небольшие группы волокон. Перимизий – покрывает более крупные комплексы мышечных волокон и мышечные пучки.
В мышце как органе есть соединительная ткань Эндомизий – тонкая соединительная ткань, окружает каждое мышечное волокно и небольшие группы волокон. Перимизий – покрывает более крупные комплексы мышечных волокон и мышечные пучки.
 Значение эндомизия и перимизия 1. Через эндомизиум и перимизиум к мышечному волокну подходят сосуды и нервы. Образуют строму органа; 2. Мышечные волокна формируются в пучки, пучки в мышцу; 3. Так как эндомизий сращен с сарколеммой мышечного волокна, следовательно, сокращающее мышечное волокно может растягиваться лишь до определенного предела
Значение эндомизия и перимизия 1. Через эндомизиум и перимизиум к мышечному волокну подходят сосуды и нервы. Образуют строму органа; 2. Мышечные волокна формируются в пучки, пучки в мышцу; 3. Так как эндомизий сращен с сарколеммой мышечного волокна, следовательно, сокращающее мышечное волокно может растягиваться лишь до определенного предела
 Миофибриллы в волокне окружены оболочкой – сарколеммой, и погружены в специальную среду – саркоплазму. В зависимости от содержания пигмента и кислорода волокна делятся на белые и красные. Белые волокна –анаэробы, содержат больше миофибрилл, меньше саркоплазмы. Запускаются быстро, но долго работать не могут. Пример: грудинно-ключичнососцевидная, икроножная мышцы. Красные волокна – волокна толстые. Много миоглобина в саркоплазме и цитохрома в митохондриях, но меньше миофибрилл. Медленно запускаются, но работают долго. Пример: мышцы спины, диафрагма.
Миофибриллы в волокне окружены оболочкой – сарколеммой, и погружены в специальную среду – саркоплазму. В зависимости от содержания пигмента и кислорода волокна делятся на белые и красные. Белые волокна –анаэробы, содержат больше миофибрилл, меньше саркоплазмы. Запускаются быстро, но долго работать не могут. Пример: грудинно-ключичнососцевидная, икроножная мышцы. Красные волокна – волокна толстые. Много миоглобина в саркоплазме и цитохрома в митохондриях, но меньше миофибрилл. Медленно запускаются, но работают долго. Пример: мышцы спины, диафрагма.
 Каждая мышца имеет сеть кровеносных сосудов. Сокращения мышцы способствует току крови. В расслабленной неработающей мышце большая часть кровеносных капилляров закрыта для тока крови. При сокращении мышцы все кровеносные капилляры сразу открываются.
Каждая мышца имеет сеть кровеносных сосудов. Сокращения мышцы способствует току крови. В расслабленной неработающей мышце большая часть кровеносных капилляров закрыта для тока крови. При сокращении мышцы все кровеносные капилляры сразу открываются.

 Строение мышцы Каждая мышца одним концом соединяется с одной костью (начало мышцы), а другим – с другой (прикрепление мышцы). В мышце различают: головку брюшко хвостовую часть.
Строение мышцы Каждая мышца одним концом соединяется с одной костью (начало мышцы), а другим – с другой (прикрепление мышцы). В мышце различают: головку брюшко хвостовую часть.
 К каждому мышечному волокну подходят двигательные нервные волокна и отходят чувствительные нервные волокна Количество нервных окончаний в мышце зависит от степени функциональной активности мышц.
К каждому мышечному волокну подходят двигательные нервные волокна и отходят чувствительные нервные волокна Количество нервных окончаний в мышце зависит от степени функциональной активности мышц.

 Каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется моторной единицей. Характерно, что мышечные волокна, принадлежащие к одной моторной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется моторной единицей. Характерно, что мышечные волокна, принадлежащие к одной моторной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.


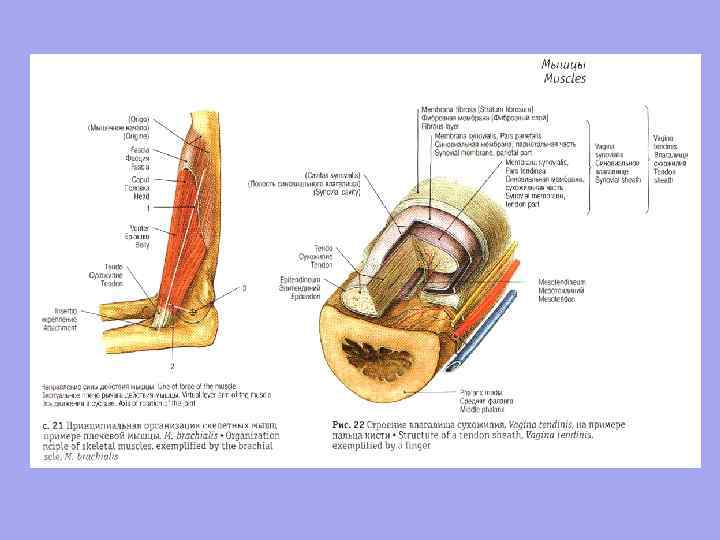
 Сухожилие – это плотный волокнистый соединительно-тканный тяж, которым мышца начинается или прикрепляется к скелету
Сухожилие – это плотный волокнистый соединительно-тканный тяж, которым мышца начинается или прикрепляется к скелету
 перитенониум коллагеновые волокна IV типа эндотенониум Коллагеновые волокна сухожилия, переплетающиеся с коллагеновыми волокнами надкостницы, вплетаются в основное вещество костной ткани, образуя на костях гребни, бугорки, бугристости, впадины, вдавления.
перитенониум коллагеновые волокна IV типа эндотенониум Коллагеновые волокна сухожилия, переплетающиеся с коллагеновыми волокнами надкостницы, вплетаются в основное вещество костной ткани, образуя на костях гребни, бугорки, бугристости, впадины, вдавления.
 Фасции – это соединительнотканные коллагенные волокна с небольшой примесью эластических волокон Поверхностная височная фасция Глубокие фасции бедра
Фасции – это соединительнотканные коллагенные волокна с небольшой примесью эластических волокон Поверхностная височная фасция Глубокие фасции бедра
 1. 2. 3. 4. 5. Фасции отделяют мышцы от кожи и устраняют смещение кожи при движениях сокращающихся мышц. Фасции экономят силу сокращения мышц, устраняя трение между мышцами во время сокращения. Фасции растягивают крупные вены при натяжении, в результате чего кровь с периферии «присасывается» в эти вены. Фасции имеют значение как барьеры, препятствующие распространению инфекции и опухолей. Во время операций фасции помогают определять расположение мышц, кровеносных сосудов, внутренностей.
1. 2. 3. 4. 5. Фасции отделяют мышцы от кожи и устраняют смещение кожи при движениях сокращающихся мышц. Фасции экономят силу сокращения мышц, устраняя трение между мышцами во время сокращения. Фасции растягивают крупные вены при натяжении, в результате чего кровь с периферии «присасывается» в эти вены. Фасции имеют значение как барьеры, препятствующие распространению инфекции и опухолей. Во время операций фасции помогают определять расположение мышц, кровеносных сосудов, внутренностей.
 Классификация мышц Скелетные мышцы разнообразны по форме, структуре, положению относительно осей суставов и т. д. , поэтому классифицируются по-разному.
Классификация мышц Скелетные мышцы разнообразны по форме, структуре, положению относительно осей суставов и т. д. , поэтому классифицируются по-разному.

 III. По функциональной особенности Статические (сильные) – короткое брюшко и длинное сухожилие. Мышцы работают с большей силой, но с меньшим размахом движения. Динамические (ловкие) – длинные мышечные пучки, сухожилия короткие. Мышцы работают с меньшей силой, но производят движения большего размаха
III. По функциональной особенности Статические (сильные) – короткое брюшко и длинное сухожилие. Мышцы работают с большей силой, но с меньшим размахом движения. Динамические (ловкие) – длинные мышечные пучки, сухожилия короткие. Мышцы работают с меньшей силой, но производят движения большего размаха
 Вспомогательный аппарат мышц Скелетные мышцы имеют вспомогательный аппарат, облегчающий их функционирование. n n n Фасции; Костно-фасциальные влагалища; Синовиальные сумки; Синовиальные влагалища сухожилий; Мышечные блоки; Сесамовидные кости.
Вспомогательный аппарат мышц Скелетные мышцы имеют вспомогательный аппарат, облегчающий их функционирование. n n n Фасции; Костно-фасциальные влагалища; Синовиальные сумки; Синовиальные влагалища сухожилий; Мышечные блоки; Сесамовидные кости.
 Аномалии развития мышц Встречаются очень часто и делятся на три группы: 1. Отсутствие какой-либо мышцы; 2. Наличие дополнительной мышцы, которой нет в природе. 3. Дополнительные пучки имеющейся мышцы.
Аномалии развития мышц Встречаются очень часто и делятся на три группы: 1. Отсутствие какой-либо мышцы; 2. Наличие дополнительной мышцы, которой нет в природе. 3. Дополнительные пучки имеющейся мышцы.
 Пороки развития Недоразвитие грудинно-ключично-сосцевидной мышцы – Кривошея Недоразвитие диафрагмы. Причина появление диафрагмальных грыж. Недоразвитие дельтовидной и трапециевидной мышц – Деформация плечевого пояса и плеча
Пороки развития Недоразвитие грудинно-ключично-сосцевидной мышцы – Кривошея Недоразвитие диафрагмы. Причина появление диафрагмальных грыж. Недоразвитие дельтовидной и трапециевидной мышц – Деформация плечевого пояса и плеча


 I. По форме: Веретенообразные; Лентовидные; Плоские широкие; Зубчатые; Длинные; n n n Квадратные; Треуголные; Круглые; Дельтовидная; Камбаловидная и т. д.
I. По форме: Веретенообразные; Лентовидные; Плоские широкие; Зубчатые; Длинные; n n n Квадратные; Треуголные; Круглые; Дельтовидная; Камбаловидная и т. д.



 II. По направлению мышечных волокон С прямыми параллельными волокнами; С поперечными; С круговыми; Перистые: A. Одноперистые; Двуперистые; C. Многоперистые. B.
II. По направлению мышечных волокон С прямыми параллельными волокнами; С поперечными; С круговыми; Перистые: A. Одноперистые; Двуперистые; C. Многоперистые. B.

 IV. По функции: Приводящие; Отводящие; Сгибающие; Разгибающие; Пронаторы; n n Супинаторы; Напрягающие; Мышцы – синергисты; Мышцы – антагонисты.
IV. По функции: Приводящие; Отводящие; Сгибающие; Разгибающие; Пронаторы; n n Супинаторы; Напрягающие; Мышцы – синергисты; Мышцы – антагонисты.
 V. По отношению к суставу: Односуставные; Двусуставные; Многосуставные.
V. По отношению к суставу: Односуставные; Двусуставные; Многосуставные.
Задачи изучения данного раздела:
- раздел помогает понять пути эволюции органического мира, постепенного усложнения не только структурных , но и физиологических, и биохимических механизмов, которое привело к возникновению современных черепных хордовых, в том числе, приматов и человека;
- в процессе сравнения удается наглядно убедиться в физиологической обусловленности появления или усложнения определенных структур, а также их исчезновения, т.о., возможно лучше понять строение человеческого тела;
- помогает осмыслить эмбриологию человека, т.к. согласно основному биогенетическому закону, сформулированному в 1866 году Геккелем и Мюллером: онтогенез есть краткое и быстрое повторение филогенеза.
Поэтому для человека в эмбриогенезе характерно появление структур, функций, биохимических реакций, которые были у его хордовых предков. Такие повторения называются рекапитуляциями . В онтогенезе человека обычно не проявляются структуры взрослых рыб, амфибий и рептилий, а только их личинок. Причем ранние стадии развития рекапитулируют более полно, чем поздние.
Но эволюционное развитие не является простым повторением черт строения предков, а предполагает изменения в ходе формирования органов и систем . Это:
- ценогенезы - приспособления зародыша к иной среде обитания, которые утрачиваются у взрослых форм, например, жабры у головастика; провизорные органы амниот;
- филэмбриогенезы - изменения развития, имеющие адаптивное значение:
а) анаболия (надставка) - добавление дополнительных стадий, например, мышечная диафрагма млекопитающих, перья птиц;
б) девиация - отклонение от программы на средних стадиях морфогенеза, например, формирование слуховых косточек, появление альвеолярного строения легких вместо ячеистого; биогенетический закон выполняется в этих случаях частично;
в) архаллаксис - отклонение онтогенеза в самом начале, когда биогенетический закон не выполняется, например, закладка волоса начинается с утолщения эпидермиса, которое погружается в кориум;
г) гетерохрония - изменение времени закладки структуры, например, ранняя закладка амниона плацентарных; и гетеротопия - изменение места закладки структуры, например, закладка плечевого пояса человека на уровне III-IV шейных позвонков, который затем перемещается на уровень I-II грудных позвонков.
При нарушениях эмбриогенеза у взрослых людей могут появиться признаки, которые присутствовали у их далеких предков - атавизмы . Если они снижают жизнеспособность, то называются атавистическими (или анцестрастральными от anĉetre) пороками . Атавизмы возникают из-за:
Недоразвития органов (гипоплазия диафрагмы, расщепление твердого неба - «волчья пасть», и др.);
Персистирования (сохранения) эмбриональных структур (незаращение боталлова протока, боковые свищи шеи, свищи пупка и др.);
Нарушения перемещения органов в онтогенезе (тазовое расположение органов, крипторхизм и др.).
Рассмотрим местоположение хордовых в системе животного мира и их классификацию .
Тип Хордовые объединяет многоклеточных вторичноротых животных различных по уровню организации. При этом абсолютно все хордовые, хотя бы на одной из стадий развития, имеют три признака, отличающих их от других животных:
- осевой комплекс органов, в основе которого лежит внутренний скелет - хорда, над ней - нервная трубка, под ней - пищеварительная трубка;
- глотка пронизана глоточными щелями, которые у высших хордовых поддерживаются висцеральными дугами;
- хвостовой отдел.
Дополнительно можно отметить наличие венозной системы воротной вены (кровь от желудочно-кишечного тракта проходит через печень) и строение покровов из эпидермиса и дермы, но похожее строение имеют покровы иглокожих.
Тип Хордовые включает четыре подтипа:
I.Полухордовые (Hemichordata) - баляноглосс, птиходера;
- II. Личиночно-хордовые (Urochordata) - асцидии;
- III. Бесчерепные (Acrania) - ланцетники;
- IV. Позвоночные (Vertebrata), которые включают несколько классов:
Группа Anamnia (низшие позвоночные)
- Круглоротые (Cyclostomata) - миноги, миксины - не имеют челюстей;
- надкласс Рыбы (Pisces):
а. класс Хрящевые и панцирные рыбы (Chondrichthyes) - акулы, скаты, химеры;
б. класс Костные рыбы (Osteichthyes) - остальные рыбы;
- Земноводные (Amphibia):
а. Хвостатые - тритоны, саламандры;
б. Бесхвостые - лягушки, жабы, квакши;
в. Безногие - червяги;
Группа Amniota (высшие позвоночные)
- Пресмыкающиеся (Reptilia):
а. Чешуйчатые - ящерицы, змеи;
б. Черепахи;
в. Крокодилы;
г. Клювоголовые - гаттерия;
- Птицы (Aves) - боковая ветвь эволюции, имеют множество специфических приспособлений
а. Бескилевые - страусы;
б. Пингвины;
в. Килевые - остальные птицы;
- Млекопитающие (Mammalia):
а. Яйцекладущие - утконос, ехидна;
б. Сумчатые - кенгуру;
в. Плацентарные - остальные звери и человек.
ФИЛОГЕНЕЗ НАРУЖНЫХ ПОКРОВОВ
Основные функции покровов - отграничение и защита многоклеточного организма. Покровы позвоночных приобретают многочисленные дополнительные функции. У хордовых покровы состоят из:
Эпидермиса, развивающегося из эктодермы;
Дермы, развивающейся из мезодермы.
Направления эволюции следующие:
Однослойный эпидермис (ланцетник) становится многослойным (у позвоночных), слои дифференцируются и ороговевают снаружи; появляются производные эпидермиса - роговые чешуи, когти, волосы и т.д.
Тонкая, студенистая соединительная ткань дермы утолщается, приобретает прочность за счет увеличения количества волокон.
Эпидермис и дерма имеют многочисленные производные: хроматофоры, чешую, роговые щитки, панцири, пластинки, клюв, когти, перья, волосы, рога, копыта. У всех хордовых есть железы. У низших - это одноклеточные слизистые железы, начиная с рыб, появляются ядовитые железы, затем сальные (у птиц), у млекопитающих добавляются млечные и потовые железы.
У человека могут быть следующие нарушения онтогенеза покровов: альбинизм (полная) и витилиго (неполная) депигментация кожи, алопеция (отсутствие) и гипертрихоз (избыточный рост) волос, гиперкератоз и пахионихия - утолщение соответственно эпидермиса и ногтей, полителия и полимастия - увеличение числа соответственно сосков и млечных желез.
ФИЛОГЕНЕЗ СКЕЛЕТА
Низшие хордовые живут в воде, высшие - преимущественно на суше. Перемещение из более плотной водной среды в воздушную привело к значительному усложнению скелета и мускулатуры, в частности, усложнился скелет конечностей и их поясов, увеличилось количество специализированных мышц, обслуживающих конечности. Основные функции скелета: опора, основа для прикрепления мышц, участие в минеральном обмене.
В основе осевого скелета лежит хорда, затем вокруг хорды сформировались позвонки, они дифференцировались, увеличивалось количество отделов позвоночника, у человека в связи с прямохождением сформировались изгибы - 2 кифоза и 2 лордоза, а остатки хорды сохранились только в центральных участках межпозвоночных дисков.
Таким образом, у рыб имеются два отдела позвоночника:
Туловищный (туловищные позвонки сочленены с ребрами);
Хвостовой;
у амфибий :
Добавляются шейный и крестцовый отделы (по 1 позвонку);
у рептилий :
Шейный (8 позвонков, I - атлант, II - аксис);
Грудной (сочленены с ребрами, некоторые из которых спереди сочленены с грудиной);
Поясничный, крестцовый (содержат большее количество позвонков);
Хвостовой;
у птиц :
Шейный (до 25 позвонков);
Грудной;
Поясничный крестцовый и часть позвонков хвостового прочно соединены, образуя мощный крестец;
у млекопитающих :
Шейный (7 позвонков);
Грудной (9-24 позвонка);
Поясничный (3-9 позвонков);
Крестцовый (позвонки срастаются);
Хвостовой (копчиковый).
У человека встречаются различные дефекты формирования позвоночного канала, через которые могут выходить спинномозговые грыжи, а также формирование патологических изгибов, в том числе боковых (сколиозы), может сохраняться хвостовой отдел.
Скелет головы делят на мозговой и висцеральный череп. В процессе эволюции шло уменьшение числа костей мозгового черепа, замена хрящевых частей на костные, уменьшение количества подвижных элементов. Число жаберных дуг висцерального скелета также прогрессивно уменьшалось. I и II жаберные дуги видоизменились и дали начало челюстям, костям лицевого черепа, слуховым костям, хрящам гортани.
У рыб I (челюстная, состоит из небно-квадратного и меккелева хрящей) дуга непосредственно соединяется со II дугой (состоит из гиомандибулярного хряща и гиоида). Такой тип соединения мозгового и лицевого черепа называется гиостильным (т.к. II дуга соединяется с гиомандибулярным хрящом). Начиная с земноводных, I дуга своим небно-квадратным хрящом срастается с основанием мозгового черепа. Такой тип соединения называется аутостильным . Из гиомандибулярного хряща формируется слуховая кость столбик, из которой в дальнейшем формируется стремечко. При нарушениях онтогенеза у человека может быть только одна эта слуховая кость (как у амфибий и рептилий) вместо трех.
Рассмотрим строение органа слуха у различных классов позвоночных. У рыб имеется только внутреннее ухо, включающее перепончатый лабиринт с отолитами, полукружные каналы и зачаток улитки.
У амфибий имеется внутреннее ухо такого же строенияи среднее ухо с одной слуховой костью и барабанной перепонкой.
У рептилий внутреннее ухо состоит из улитки и полукружных каналов, среднее - такое же как у амфибий, есть зачаток наружного слухового прохода.
У птиц хорошо сформирован наружный слуховой проход.
У млекопитающих внутренее ухо включает улитку 2,5 оборота, полукружные каналы, мешочек, маточку; среднее ухо - барабанную полость, в которой находятся слуховые кости - стремечко (остаток гиомандибулярного хряща), наковальня (остаток небно-квадратного хряща), молоточек (остаток меккелева хряща), имеется барабанная перепонка, наружное включает наружный слуховой проход и ушную раковину.
Таким образом, в висцеральном скелете в процессе эволюции хрящевые челюсти сменялись костными, изменился тип крепления челюстей, изменилась функция элементов висцеральных дуг.
В скелете конечностей сформировались пояса с подвижно прикрепленным скелетом свободных конечностей; сформировалась пятипалая конечность наземного типа; уменьшилось число костей в дистальных отделах; удлинялись проксимальные и укорачивались дистальные отделы конечностей.
У человека могут быть следующие пороки: полидактилия и полифалангия (увеличение числа пальцев и фаланг соответственно); перемещение пояса верхних конечностей на уровень I-II грудных позвонков - болезнь Шпренгеля ; врожденные аномалии ребер и деформации грудной клетки - воронководная , килевидная и др.
ФИЛОГЕНЕЗ МЫШЦ
Мускулатура выполняет функцию движения и делится на:
Соматическую, которая развивается из миотомов и иннервируется спинномозговыми нервами (поперечно-полосатая);
Висцеральную - развивается из других участков мезодермы, иннервируется вегетативными нервами, может быть поперечно-полосатой и гладкой.
У низших хордовых висцеральная мускулатура располагается в области жаберных дуг, постепенно из нее формируются жевательные мышцы, мышцы глотки, затем мышцы шеи и плечевого пояса, а также мимическая мускулатура. Мышцы шеи и лица имеют сложную иннервацию, что связано с этапами филогенеза. Соматическая мускулатура у низших хордовых расположена в виде миомеров, затем формируются мускулатура конечностей, более разнообразной становится форма, размеры и функции мышц, усиливается их иннервация, особенно свободных конечностей у приматов. Мышцы туловища в процессе эволюции редуцируются.